Interspecific Hybridization and Introgression Influence Biodiversity—Based on Genetic Diversity of Central European Viola epipsila-V. palustris Complex

 , , , ,
, , , ,
Abstract
:
1. Introduction
2. Material and Methods
2.1. Plant Material Collection
2.2. Morphological Analyses
2.3. DNA Extraction and ISSR Analysis
2.4. Genome Size Assessment
2.5. Sequencing of cpDNA
3. Results
3.1. Variability of Morphological Features
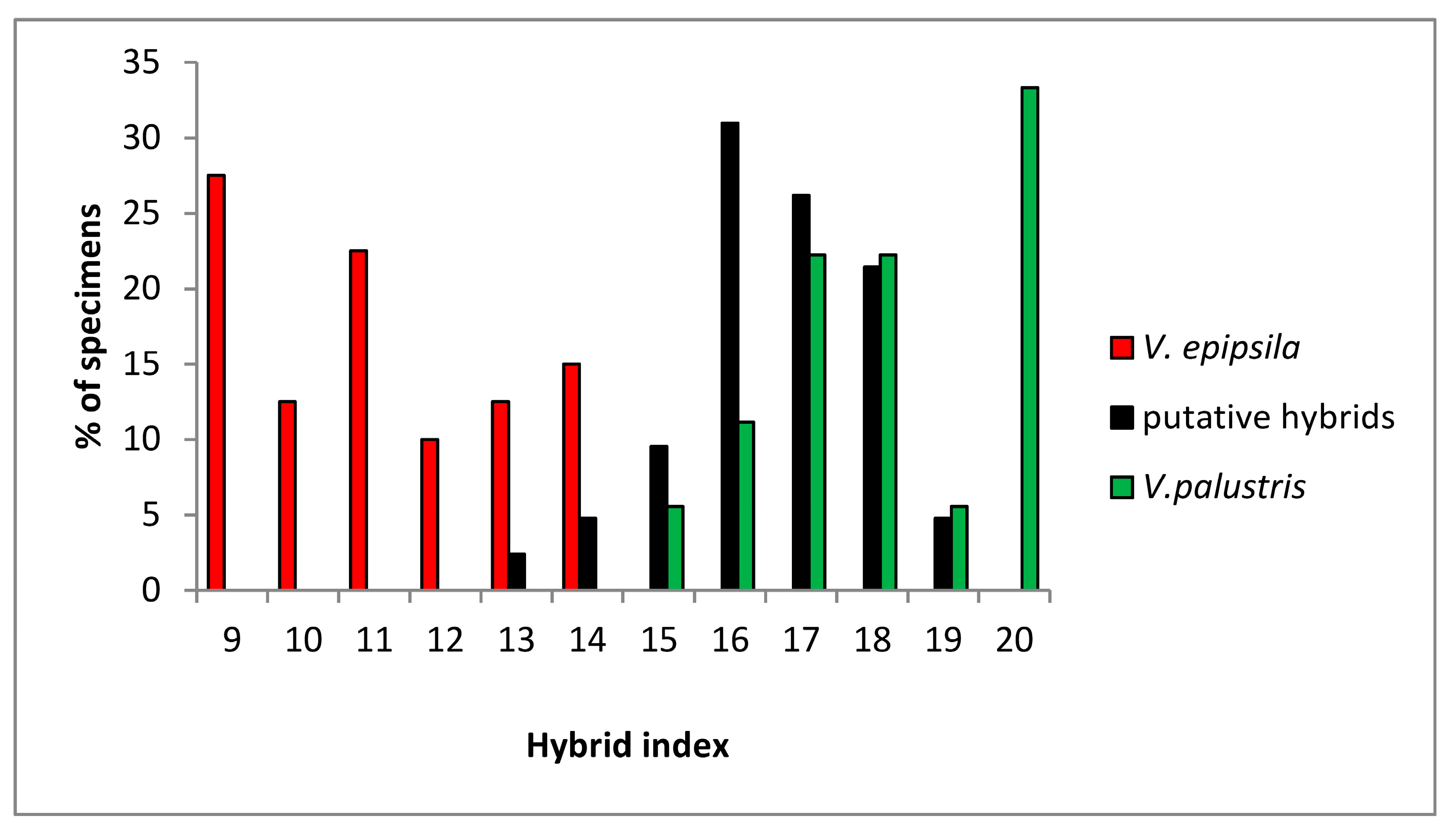
3.2. Genetic Diversity of V. epipsila-V. palustis Complex Increased by Hybrids and Introgressants
3.3. Genome Size as a Good Marker to Distinguish Parental Species and Their F1 Hybrids
3.4. Viola epipsila Is Maternal Species of Hybrids Based on Non-Coding cpDNA Sequences
4. Discussion
4.1. Difficulties in the Identification of V. epipsila, V. palustris and their Putative Hybrids in the Field
4.2. Is There a Risk of Extinction of V. epipsila in Central Europe?
4.3. Local Sympatric Populations of V. epipsila and V. palustris as a Good Model for Studying the Hybridization Effect on Population Structure
4.4. Low Intrapopulation Genetic Diversity of the V. epipsila-V. palustris Complex and Unidirectional Crosses
4.5. Lack of Intraspecific Variation in Genome Size
5. Conclusions and Further Perspectives
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pauls, S.U.; Nowak, C.; Bálint, M.; Pfenninger, M. The impact of global climate change on genetic diversity within populations and species. Mol. Ecol. 2013, 22, 925–946. [Google Scholar] [CrossRef] [PubMed]
- Schierenbeck, K.A. Population-level genetic variation and climate change in a biodiversity hotspot. Ann. Bot. 2017, 119, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Wróblewska, A.; Mirski, P. From past to future: Impact of climate change on range shifts and genetic diversity patterns of circumboreal plants. Reg. Environ. Chang. 2018, 18, 409–424. [Google Scholar] [CrossRef] [Green Version]
- Arnold, M.J. Natural Hybridization and Evolution; Oxford University Press: New York, NY, USA, 1997. [Google Scholar]
- Rieseberg, L.H. Hybrid origins of plant species. Annu. Rev. Ecol. Syst. 1997, 28, 359–389. [Google Scholar] [CrossRef] [Green Version]
- Wolf, D.E.; Takebayashi, N.; Rieseberg, L.H. Predicting the risk of extinction through hybridization. Conserv. Biol. 2001, 15, 1039–1053. [Google Scholar] [CrossRef]
- Mallet, J. Hybrid speciation. Nature 2007, 446, 278–283. [Google Scholar] [CrossRef]
- Alix, K.; Gérard, P.R.; Schwarzacher, T.; Heslop-Harrison, J.S. Polyploidy and interspecific hybridization: Partners for adaptation, speciation and evolution in plants. Ann. Bot. 2017, 120, 183–194. [Google Scholar] [CrossRef]
- Rieseberg, L.H.; Ellstrand, N.C. What can molecular and morphological markers tells us about plant hybridization? CRC Crit. Rev. Plant Sci. 1993, 12, 213–241. [Google Scholar] [CrossRef]
- Twyford, A.D.; Ennos, R.A. Next-generation hybridization and introgression. Heredity 2012, 108, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Goulet, B.E.; Roda, F.; Hopkins, R. Hybridization in plants: Old ideas, new techniques. Plant Physiol. 2017, 173, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Hegi, G. Viola L. In Illustrierte Flora von Mitteleuropa; J.F. Lehmann: München, Germany, 1925; Volume 5, pp. 586–656. [Google Scholar]
- Komarov, V.L. Flora URSS; Academia Scientiarum URSS: Leningrad, Russia, 1949; Volume 15, p. 742. [Google Scholar]
- Sorsa, M. Cytological and evolutionary studies on Palustres violets. Madroño 1968, 19, 165–179. [Google Scholar]
- Valentine, D.H.; Merxmüller, H.; Schmidt, A. Viola L. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Eds.; Cambridge University Press: Cambridge, UK, 1968; Volume 2, pp. 270–282. [Google Scholar]
- Meusel, H.; Jäger, E.; Weinert, E. Vergleichende Chorologie der Zentraleuropäischen Flora; G. Fischer: Jena, Germany, 1978; Volume 2, p. 418. [Google Scholar]
- Hulten, E.; Fries, M. Atlas of North European Vascular Plants North of the Tropic of Cancer; Koeltz Scientific Books: Königstein, Germany, 1986; Volume 3, pp. 969–1172. [Google Scholar]
- Karlsson, T.; Marcussen, T. Violaceae. In Flora Nordica; Jonsell, B., Karlsson, T., Eds.; The Swedish Museum of Natural History: Stockholm, Sweden, 2010; Volume 6, pp. 12–52. [Google Scholar]
- Anderson, J.P. Flora of Alaska and Adjacent Parts of Canada; University of Iowa Press: Iowa, IA, USA, 1959; p. 543. [Google Scholar]
- Polunin, N. Circumpolar Arctic Flora; Clarendon Press: Oxford, UK, 1959; p. 514. [Google Scholar]
- Löve, A.; Löve, E. Cytotaxonomical Atlas of the Arctic Flora; Cramer: Vaduz, Liechtenstein, 1975; p. 598. [Google Scholar]
- Tolmatchev, A.I.; Jurtsev, B.A. Flora Arctica URSS; Nauka: Leningrad, Russia, 1980; Volume 8, p. 331. [Google Scholar]
- Kuta, E. Biosystematic studies on the genus Viola L.; section Plagiostigma Godr. I. Karyological analysis of V. epipsila Ledeb.; V. palustris L. and their hybrids from Poland. Acta Biol. Cracov. Bot. 1989, 31, 29–44. [Google Scholar]
- Kuta, E. Biosystematic studies on the genus Viola L.; section Plagiostigma Godr. II. Embryological analysis of V. epipsila Ledeb.; V. palustris L. and their hybrids from Poland. Acta Biol. Cracov. Bot. 1989, 31, 45–62. [Google Scholar]
- Kuta, E. Biosystematic studies on Viola sect. Plagiostigma: III. Biometrical analysis of the Polish populations of V. epipsila, V. palustris and their spontaneous hybrids. Fragm. Florist. Geobot. 1991, 35, 5–34. [Google Scholar]
- Loos, G.H. Taxonomische Neukombinationen zur Flora Mittel-und Osteuropas, insbesondere Nordrhein-Westfalens. Online-Veröff. Boch. Bot. Ver. 2010, 2, 1–20. [Google Scholar]
- Anderson, E. Introgressive Hybridization; Wiley: New York, NY, USA, 1949. [Google Scholar]
- Index of/src/base/R-3. Available online: https://cran.r-project.org/src/base/R-3/ (accessed on 20 May 2020).
- Gupta, M.; Chyi, Y.S.; Romero-Severson, J.; Owen, J.L. Amplification of DNA markers from evolutionary diverse genomes using single primers of simple-sequence repeats. Theor. Appl. Genet. 1994, 89, 998–1006. [Google Scholar] [CrossRef] [PubMed]
- Stepansky, A.; Kovalski, I.; Perl-Treves, R. Intraspecific classifcation of melons (Cucumis melo L.) in view of their phenotypic and molecular variation. Plant Syst. Evol. 1999, 217, 313–332. [Google Scholar] [CrossRef]
- Yeh, F.C.; Yang, R.C.; Boyle, T. POPGENE. In Microsoft Windows Based Freeware for Population Genetic Analysis. Release 1.31; University of Alberta: Edmonton, AB, Canada, 1999; Available online: https://sites.ualberta.ca/~fyeh/popgene.html (accessed on 18 August 2020).
- Schlüter, P.M.; Harris, S.A. Analysis of multilocus fingerprinting data sets containing missing data. Mol. Ecol. Notes 2006, 6, 569–572. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Halverson, K.; Heard, S.B.; Nason, J.D.; Stireman, J.O. Origins, distribution, and local co-occurrence of polyploid cytotypes in Solidago altissima (Asteraceae). Am. J. Bot. 2008, 95, 50–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnier, J.; Buerki, S.; Arrigo, N.; KÜpfer, P.; Alvarez, N. Genetic structure and evolution of Alpine polyploid complexes: Ranunculus kuepferi (Ranunculaceae) as a case study. Mol. Ecol. 2009, 18, 3730–3744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampson, J.F.; Byrne, M. Genetic diversity and multiple origins of polyploid Atriplex nummularia Lindl. (Chenopodiaceae). Biol. J. Linn. Soc. Lond. 2012, 105, 218–230. [Google Scholar] [CrossRef] [Green Version]
- Klie, M.; Menz, I.; Linde, M.; Debener, T. Lack of structure in the gene pool of the highly polyploidy ornamental chrysanthemum. Mol. Breed. 2013, 32, 339–348. [Google Scholar] [CrossRef]
- Laport, R.G.; Minckley, R.L.; Ramsey, J. Ecological distributions, phenological isolation, and genetic structure in sympatric and parapatric populations of the Larrea tridentata polyploid complex. Am. J. Bot. 2016, 103, 1358–1374. [Google Scholar] [CrossRef] [Green Version]
- Kuta, E.; Jędrzejczyk-Korycińska, M.; Cieślak, E.; Rostański, A.; Szczepaniak, M.; Migdałek, G.; Wasowicz, P.; Suda, J.; Combik, M.; Slomka, A. Morphological versus genetic diversity of Viola reichenbachiana and V. riviniana (sect. Viola, Violaceae) from soils differing in heavy metal content. Plant Biol. (Stuttg.) 2014, 16, 924–934. [Google Scholar] [CrossRef]
- Migdałek, G.; Nowak, J.; Saluga, M.; Cieslak, E.; Szczepaniak, M.; Ronikier, M.; Marcussen, T.; Slomka, A.; Kuta, E. No evidence of contemporary interploidy gene flow between the closely related European woodland violets Viola reichenbachiana and V. riviniana (sect. Viola, Violaceae). Plant Biol. (Stuttg.) 2017, 19, 542–551. [Google Scholar] [CrossRef]
- Besnier, F.; Glover, K.A. ParallelStructure: AR package to distribute parallel runs of the population genetics program STRUCTURE on multi-core computers. PLoS ONE 2013, 8, e7065. [Google Scholar] [CrossRef] [Green Version]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. Clumpak: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef] [Green Version]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.C.; Thompson, E.A. A model-based method for identifying species hybrids using multilocus genetic data. Genetics 2002, 160, 1217–1229. [Google Scholar]
- Thiem, B.; Sliwinska, E. Flow cytometric analysis of nuclear DNA content in cloudberry (Rubus chamaemorus L.) In Vitro cultures. Plant Sci. 2003, 164, 129–134. [Google Scholar] [CrossRef]
- Doležel, J.; Sgorbati, S.; Lucretti, S. Comparison of three DNA fluorochromes for flow cytometric estimation of nuclear DNA content in plants. Physiol. Plant 1992, 85, 625–631. [Google Scholar] [CrossRef]
- Hamilton, M.B. Primer Notes: Four primer pairs for the amplification of chloroplast intergenic regions with intraspecific variation. Mol. Ecol. 1999, 8, 513–525. [Google Scholar]
- Borsch, T.; Quandt, D. Mutational dynamics and phylogenetic utility of noncoding chloroplast DNA. Plant Syst. Evol. 2009, 282, 169–199. [Google Scholar] [CrossRef]
- Cennamo, P.; Guacchio, E.D.; Jury, S.L.; Caputo, P. Molecular markers in Viola L. subsect. Viola: Application and taxonomic implications for the identification of dubious herbarium specimens. Plant Biosyst. 2011, 145, 306–323. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Simmons, M.P.; Ochoterena, H. Gaps as characters in sequence-based phylogenetic analyses. Syst. Biol. 2000, 49, 369–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, K. SeqState—Primer design and sequence statistics for phylogenetic DNA data sets. Appl. Bioinform. 2005, 4, 65–69. [Google Scholar] [CrossRef]
- Marcussen, T.; Jakobsen, K.S.; Danihelka, J.; Ballard, H.E.; Blaxland, K.; Brysting, A.K.; Oxelman, B. Inferring species networks from gene trees in high-polyploid North American and Hawaiian violets (Viola, Violaceae). Syst. Biol. 2012, 61, 107–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcussen, T.; Heier, L.; Brysting, A.K.; Oxelman, B.; Jakobsen, K.S. From gene trees to a dated allopolyploid network: Insights from the angiosperm genus Viola (Violaceae). Syst. Biol. 2015, 64, 84–101. [Google Scholar] [CrossRef] [Green Version]
- Stace, C.A. Hybridization and the Flora of the British Isles; Academic Press: London, UK, 1975; p. 626. [Google Scholar]
- Ellstrand, N.C.; Whitkus, R.; Rieseberg, L.H. Distribution of spontaneous plant hybrids. Proc. Natl. Acad. Sci. USA 1996, 93, 5090–5093. [Google Scholar] [CrossRef] [Green Version]
- Heiser, C.B. Introgression re-examined. Bot. Rev. 1973, 39, 347–366. [Google Scholar] [CrossRef]
- Brainerd, E. Hybridism in the genus Viola. Rhodora 1904, 6, 213–223. [Google Scholar] [CrossRef]
- Russell, N.H. Local introgression between Viola cucullata Ait. and V. septentrionalis Greene. Evolution 1955, 9, 436–440. [Google Scholar] [CrossRef]
- Russell, N.H.; Risser, A.C. The hybrid nature of Viola emarginata (Nuttall) Leconte. Brittonia 1960, 12, 298–305. [Google Scholar] [CrossRef]
- Moore, D.M.; Harvey, M.J. Cytogenetic relationships of Viola lactea Sm. and other west European arosulate violets. N. Phytol. 1961, 60, 85–95. [Google Scholar] [CrossRef]
- Kuta, E. Cyto-embryological studies on the species of the Viola L. genus., Nomimium Ging. section from the territory of Poland. Fragm. Florist. Geobot. 1978, 24, 23–91. [Google Scholar]
- Kuta, E. Further cyto-embryological studies on Viola L. section Viola L. Acta Biol. Cracov. Bot. 1981, 23, 69–82. [Google Scholar]
- Erben, M. The significance of hybridization on the forming of species in the genus Viola. Bocconea 1996, 5, 113–118. [Google Scholar]
- Neuffer, B.; Auge, H.; Mesch, H.; Amarell, U.; Brandl, R. Spread of violets in polluted pine forests: Morphological and molecular evidence for the ecological importance of interspecific hybridization. Mol. Ecol. 1999, 8, 365–377. [Google Scholar] [CrossRef]
- Conesa, M.À.; Mus, M.; Rosselló, J.A. Hybridization between insular endemic and widespread species of Viola in non-disturbed environments assessed by nuclear ribosomal and cpDNA sequences. Plant Syst. Evol. 2008, 273, 169–177. [Google Scholar] [CrossRef]
- van den Hof, K.; van den Berg, R.G.; Gravendeel, B. Chalcone synthase gene lineage diversification confirms allopolyploid evolutionary relationships of European rostrate violets. Mol. Biol. Evol. 2008, 25, 2099–2108. [Google Scholar] [CrossRef] [Green Version]
- Migdałek, G.; Woźniak, M.; Słomka, A.; Godzik, B.; Jędrzejczyk-Korycińsa, M.; Rostański, A.; Bothe, H.; Kuta, E. Morphological differences between violets growing at heavy metal polluted and non-polluted sites. Flora 2013, 208, 87–96. [Google Scholar] [CrossRef]
- Blaxland, K.; Ballard, H.E.; Marcussen, T. Viola pluviae sp. nov. (Violaceae), a member of subsect. Stolonosae in the Pacific Northwest region of North America. Nord. J. Bot. 2018, 36. [Google Scholar] [CrossRef]
- Brainerd, E. The natural violet hybrids of North America. Bull. Vermont Agric. Exp. Stn. 1924, 239, 1–205. [Google Scholar]
- Zabłocki, J. Dwuliścienne wolnopłatkowe: Dwuokwiatowe, rodzina: Violaceae, fiołkowate. In Flora Polska. Rośliny Naczyniowe Polski i Ziem Ościennych; Szafer, W., Ed.; Polska Akademia Umiejętności: Kraków, Poland, 1947; Volume 6, pp. 1–70. [Google Scholar]
- Bettinger, A.; Buttler, K.P.; Caspari, S.; Klotz, J.; May, R.; Metzing, D. Verbreitungsatlas der Farn-und Blütenpflanzen Deutschlands; Netzwerk Phytodiversität Deutschlands, Bundesamt für Naturschutz: Bonn, Germany, 2013; p. 834. [Google Scholar]
- Rätzel, S. Viola epipsila Ledeb. Verh. Bot. Ver. 2018, 150, 199–202. [Google Scholar]
- Kuta, E.; Pliszko, A. Viola epipsila Ledeb. In Polska Czerwona Księga Roślin, Paprotniki i Rośliny Kwiatowe; Kazimierczakowa, R., Zarzycki, K., Mirek, Z., Eds.; Instytut Ochrony Przyrody PAN: Kraków, Polska, 2014; Volume 3, pp. 329–331. [Google Scholar]
- Gompert, Z.; Buerkle, C.A. What, if anything, are hybrids: Enduring truths and challenges associated with population structure and gene flow. Evol. Appl. 2016, 9, 909–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, E. Hybridization of the habitat. Evolution 1948, 2, 1–9. [Google Scholar] [CrossRef]
- Anderson, E.; Stebbins, G.L. Hybridization as an evolutionary stimulus. Evolution 1954, 8, 378–388. [Google Scholar] [CrossRef]
- Arnold, M.L. Anderson’s and Stebbins’ prophesy comes true: Genetic exchange in fluctuating environments. Syst. Bot. 2016, 41, 4–16. [Google Scholar] [CrossRef]
- McFarlane, S.R.; Pemberton, J.M. Detecting the true extent of introgression during anthropogenic hybridization. Trends Ecol. Evol. (Amst.) 2019, 34, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Brzeziecki, B. Long-term tree and stand dynamics. In The Forests of the Strict Reserve of Białowieża National Park; Keczyński, A., Ed.; Białowieża National Park: Białowieża, Poland, 2017; pp. 94–125. [Google Scholar]
- Latałowa, M.; Zimny, M.; Jędrzejewska, B.; Samojlik, T. Białowieża Primeval Forest: A 2000-year Interplay of Environmental and Cultural Forces in Europe’s Best Preserved Temperate Woodland. In Europe’s Changing Woods and Forests: From Wildwood to Managed Landscapes; Kirby, K.J., Watkins, C., Eds.; Oxford University: Oxford, UK, 2015; pp. 243–264. [Google Scholar]
- Latałowa, M.; Zimny, M.; Pędziszewska, A.; Kupryjanowicz, M. Postglacjalna historia Puszczy Białowieskiej—roślinność, klimat i działalność człowieka. Parki Nar. Rez. Przyr. 2016, 35, 3–49. [Google Scholar]
- Cholewińska, O.; Adamowski, W.; Jaroszewicz, B. Homogenization of temperate mixed deciduous forests in Białowieża forest: Similar communities are becoming more similar. Forests 2020, 11, 545. [Google Scholar] [CrossRef]
- Sokołowski, A. Rośliny naczyniowe nadleśnictwa Zwierzyniec w Puszczy Białowieskiej. Fragm. Florist. Geobot. 1968, 14, 131–150. [Google Scholar]
- Reddy, M.P.; Sarla, N.; Siddiq, E. Inter simple sequence repeat (ISSR) polymorphism and its application in plant breeding. Euphytica 2002, 128, 9–17. [Google Scholar] [CrossRef]
- Hamrick, J.L.; Godt, M.J.W. Allozyme diversity in plant species. In Plant Population Genetics, Breeding, and Genetic Resources; Brown, A.H.D., Clegg, M.T., Kahler, A.L., Weir, B.S., Eds.; Sinauer Associates: Sunderland, MA, USA, 1990; pp. 43–63. [Google Scholar]
- Auge, H.; Neuffer, B.; Erlinghagen, F.; Grupe, R.; Brandl, R. Demographic and random amplified polymorphic DNA analyses reveal high levels of genetic diversity in a clonal violet. Mol. Ecol. 2001, 10, 1811–1819. [Google Scholar] [CrossRef]
- Cánovas, J.L.; Jimenez, J.F.; Mota, J.F.; Gomez, P.S. Genetic diversity of Viola cazorlensis Gand., an endemic species of Mediterranean dolomitic habitats: Implications for conservation. Syst. Biodivers. 2015, 13, 571–580. [Google Scholar] [CrossRef]
- Lavania, U.C. Plant speciation and polyploidy: In habitat divergence and environmental perspective. Nucleus 2020, 63, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Rieseberg, L.H.; Baird, S.J.E.; Gardner, K.A. Hybridization, introgression, and linkage evolution. Plant Mol. Biol. 2000, 42, 205–224. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Bendixsen, D.P.; Janzen, T.; Nolte, A.W.; Greik, D.; Stelkens, R. Recombining your way out of trouble: The genetic architecture of hybrid fitness under environmental stress. Mol. Biol. Evol. 2020, 37, 167–182. [Google Scholar] [CrossRef] [Green Version]
- Gramlich, S.; Wagner, N.D.; Hörandl, E. RAD-seq reveals genetic structure of the F2-generation of natural willow hybrids (Salix L.) and a great potential for interspecific introgression. BMC Plant Biol. 2018, 18, 317. [Google Scholar] [CrossRef]
- Paetzold, C.; Wood, K.R.; Eaton, D.A.R.; Wagner, W.L.; Appelhans, M.S. Phylogeny of Hawaiian Melicope (Rutaceae): RAD-seq resolves species relationships and reveals ancient introgression. Front. Plant Sci. 2019, 10, 1074. [Google Scholar] [CrossRef] [Green Version]
- Knight, C.A.; Ackerly, D.D. Variation in nuclear DNA content across environmental gradients: A quantile regression analysis. Ecol. Lett. 2002, 5, 66–76. [Google Scholar] [CrossRef] [Green Version]
- Bennetzen, J.L.; Ma, J.; Devos, K.M. Mechanisms of recent genome size variation in flowering plants. Ann. Bot. 2005, 95, 127–132. [Google Scholar] [CrossRef] [Green Version]
- Qiu, F.; Baack, E.J.; Whitney, K.D.; Bock, D.G.; Tetreault, H.M.; Rieseberg, L.H.; Ungerer, M.C. Phylogenetic trends and environmental correlates of nuclear genome size variation in Helianthus sunflowers. N. Phytol. 2019, 221, 1609–1618. [Google Scholar] [CrossRef] [Green Version]
- Greilhuber, J.; Doležel, J.; Lysák, M.A.; Bennett, M.D. The origin, evolution and proposed stabilization of the terms ‘genome size’ and ‘C-value’ to describe nuclear DNA contents. Ann. Bot. 2005, 95, 255–260. [Google Scholar] [CrossRef]
- Leitch, I.J.; Bennett, M.D. Genome downsizing in polyploid plants. Biol. J. Linn. Soc. Lond. 2004, 82, 651–663. [Google Scholar] [CrossRef] [Green Version]
- Poggio, L.; Realini, M.F.; Fourastié, M.F.; Garcìa, A.M.; González, G.E. Genome downsizing and karyotype constancy in diploid and polyploid congeners: A model of genome size variation. AoB Plants 2014, 6, plu029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baack, E.J.; Whitney, K.D.; Rieseberg, L.H. Hybridization and genome size evolution: Timing and magnitude of nuclear DNA content increases in Helianthus homoploid hybrid species. N. Phytol. 2005, 167, 623–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bashir, T.; Mishra, R.C.; Hasan, M.M.; Mohanta, T.K.; Bae, H. Effect of hybridization on somatic mutations and genomic rearrangements in plants. Int. J. Mol. Sci. 2018, 19, 3758. [Google Scholar] [CrossRef] [Green Version]
- Slazak, B.; Śliwińska, E.; Saługa, M.; Ronikier, M.; Bujak, J.; Słomka, A.; Göransson, U.; Kuta, E. Micropropagation of Viola uliginosa (Violaceae) for endangered species conservation and for somaclonal variation-enhanced cyclotide biosynthesis. Plant Cell Tissue Organ Cult. 2015, 120, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Żabicki, P.; Sliwinska, E.; Mitka, J.; Sutkowska, A.; Tuleja, M.; Migdałek, G.; Żabicka, J.; Słomka, A.; Kwiatkowska, M.; Kuta, E. Does somaclonal variation play advantageous role in conservation practice of endangered species? Comprehensive genetic studies of In Vitro propagated plantlets of Viola stagnina Kit. (Violaceae). Plant Cell Tissue Organ Cult. 2019, 136, 339–352. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | N * | Mean of Length of Sepals with Calycine Appendages (mm) (±SD) | Mean of Length of Calycine Appendages (mm) (±SD) | Mean of Length of Spur (mm) (±SD) | Mean of Length of Lower Petal with Spur (mm) (±SD) | Mean of Ratio of Leaf Length to Width (±SD) | Mean of Ratio of Distance of Bracts from Rhizome to Length of Pedicel (±SD) | Mean of Length of Flower (mm) (±SD) | Mean of Width of Flower (mm) (±SD) |
|---|---|---|---|---|---|---|---|---|---|
| V. epipsila | 40 | 6.68 a (±0.79) | 1.41 a (±0.42) | 9.70 a (±0.88) | 17.85 a (±1.73) | 0.79 a (±0.14) | 0.69 a (±0.08) | 21.55 a (±2.45) | 24.88 a (±2.84) |
| putative hybrids | 43 | 6.19 b (±0.69) | 1.19 a (±0.29) | 6.62 b (±0.80) | 13.59 b (±1.25) | 0.82 a (±0.12) | 0.49 b (±0.07) | 18.42 b (±1.40) | 20.05 b (±2.11) |
| V. palustris | 18 | 5.33 c (±0.69) | 0.89 b (±0.27) | 6.28 b (±0.57) | 11.67 c (±1.19) | 0.79 a (±0.15) | 0.40 c (±0.08) | 13.89 c (±2.49) | 15.44 c (±2.83) |
| Taxon/Pop * | N | P(%poly) | Nprt | Hj | HT | HS | GST | |
|---|---|---|---|---|---|---|---|---|
| V. epipsila | 0.095 (0.027) | 0.035 (0.004) | 0.636 | |||||
| BNP | 13 | 6 (3.59) | 7 | 4 | 0.015 (0.080) | |||
| SZ | 4 | 21 (12.57) | 7 | 1 | 0.052 (0.138) | |||
| L | 10 | 23 (13.77) | 7 | 1 | 0.036 (0.098) | |||
| Total | 27 | 55 (32.93) | 21 | 6 | 0.083 (0.155) | |||
| V. palustris | 0.083 (0.024) | 0.012 (0.001) | 0.850 | |||||
| BNP | 8 | 0 (0.00) | 1 | 1 | 0.000 (0.000) | |||
| SZ | 5 | 13 (7.78) | 1 | 0 | 0.035 (0.123) | |||
| N_NE | 9 | 0 (0.00) | 0 | 0 | 0.000 (0.000) | |||
| N_B | 14 | 4 (2.40) | 0 | 0 | 0.011 (0.070) | |||
| N_SE | 5 | 0 (0.00) | 0 | 0 | 0.000 (0.000) | |||
| BAL | 10 | 0 (0.00) | 1 | 1 | 0.000 (0.000) | |||
| KR | 21 | 1 (0.60) | 0 | 0 | 0.002 (0.024) | |||
| L | 10 | 23 (13.77) | 2 | 0 | 0.051 (0.132) | |||
| Total | 82 | 50 (29.94) | 5 | 2 | 0.074 (0.150) | |||
| hybrids | 0.101 (0.030) | 0.003 (0.000) | 0.974 | |||||
| BNP | 73 | 0 (0.00) | 5 | 5 | 0.000 (0.000) | |||
| SZ | 6 | 0 (0.00) | 0 | 0 | 0.000 (0.000) | |||
| KOZ | 15 | 6 (3.59) | 1 | 0 | 0.008 (0.042) | |||
| OST | 8 | 2 (1.20) | 0 | 0 | 0.006 (0.051) | |||
| L | 4 | 0 (0.00) | 6 | 6 | 0.000 (0.000) | |||
| Total | 106 | 45 (26.95) | 12 | 11 | 0.061 (0.130) |
| Source of Variation | df | Sum of Squares | Variance Components | Percentage of Variation | F, p-Value |
|---|---|---|---|---|---|
| V. epipsila vs. V. palustris | |||||
| among groups | 1 | 1044.50 | 26.30 | 79.18 | FCT = 0.79 P = 0.012 |
| among populations within groups | 6 | 366.19 | 5.89 | 17.72 | FSG = 0.85 a |
| within populations | 78 | 80.36 | 1.03 | 3.10 | FST = 0.97 a |
| total | 85 | 1491.05 | 33.22 | - | |
| V. epipsila vs. V. palustris vs. hybrids | |||||
| among groups | 2 | 1985.09 | 12.80 | 59.92 | FCT = 0.60 a |
| among populations within groups | 13 | 1089.63 | 7.80 | 36.53 | FSG = 0.91 a |
| within populations | 199 | 150.84 | 0.76 | 3.55 | FST = 0.96 a |
| total | 214 | 3225.56 | 21.36 | - | |
| V. epipsila | |||||
| among populations | 2 | 123.81 | 7.20 | 72.90 | FST = 0.73 a |
| within populations | 24 | 64.27 | 2.68 | 27.10 | |
| total | 26 | 188.07 | 9.88 | - | |
| V. palustris | |||||
| among populations | 4 | 242.38 | 5.39 | 94.76 | FST = 0.95 a |
| within populations | 54 | 16.10 | 0.30 | 5.24 | |
| total | 58 | 258.48 | 5.69 | - | |
| hybrids | |||||
| among populations | 5 | 545.03 | 10.08 | 98.73 | FST = 0.99 a |
| within populations | 101 | 13.08 | 0.130 | 1.27 | |
| total | 106 | 558.11 | 10.21 | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Żabicka, J.; Migdałek, G.; Słomka, A.; Sliwinska, E.; Mackiewicz, L.; Keczyński, A.; Kuta, E. Interspecific Hybridization and Introgression Influence Biodiversity—Based on Genetic Diversity of Central European Viola epipsila-V. palustris Complex. Diversity 2020, 12, 321. https://doi.org/10.3390/d12090321
Żabicka J, Migdałek G, Słomka A, Sliwinska E, Mackiewicz L, Keczyński A, Kuta E. Interspecific Hybridization and Introgression Influence Biodiversity—Based on Genetic Diversity of Central European Viola epipsila-V. palustris Complex. Diversity. 2020; 12(9):321. https://doi.org/10.3390/d12090321
Chicago/Turabian StyleŻabicka, Justyna, Grzegorz Migdałek, Aneta Słomka, Elwira Sliwinska, Leszek Mackiewicz, Andrzej Keczyński, and Elżbieta Kuta. 2020. "Interspecific Hybridization and Introgression Influence Biodiversity—Based on Genetic Diversity of Central European Viola epipsila-V. palustris Complex" Diversity 12, no. 9: 321. https://doi.org/10.3390/d12090321