Structure and Genetic Diversity of Nine Important Landraces of Capsicum Species Cultivated in the Yucatan Peninsula, Mexico


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Extraction and ISSR Amplification
2.3. Data Analysis
2.3.1. Genetic Structure Analysis
2.3.2. Genetic Relationship Analysis
2.3.3. Genetic Diversity Analysis
3. Results
3.1. Genetic Structure of Capsicum Species Landraces Cultivated in the Yucatan Peninsula
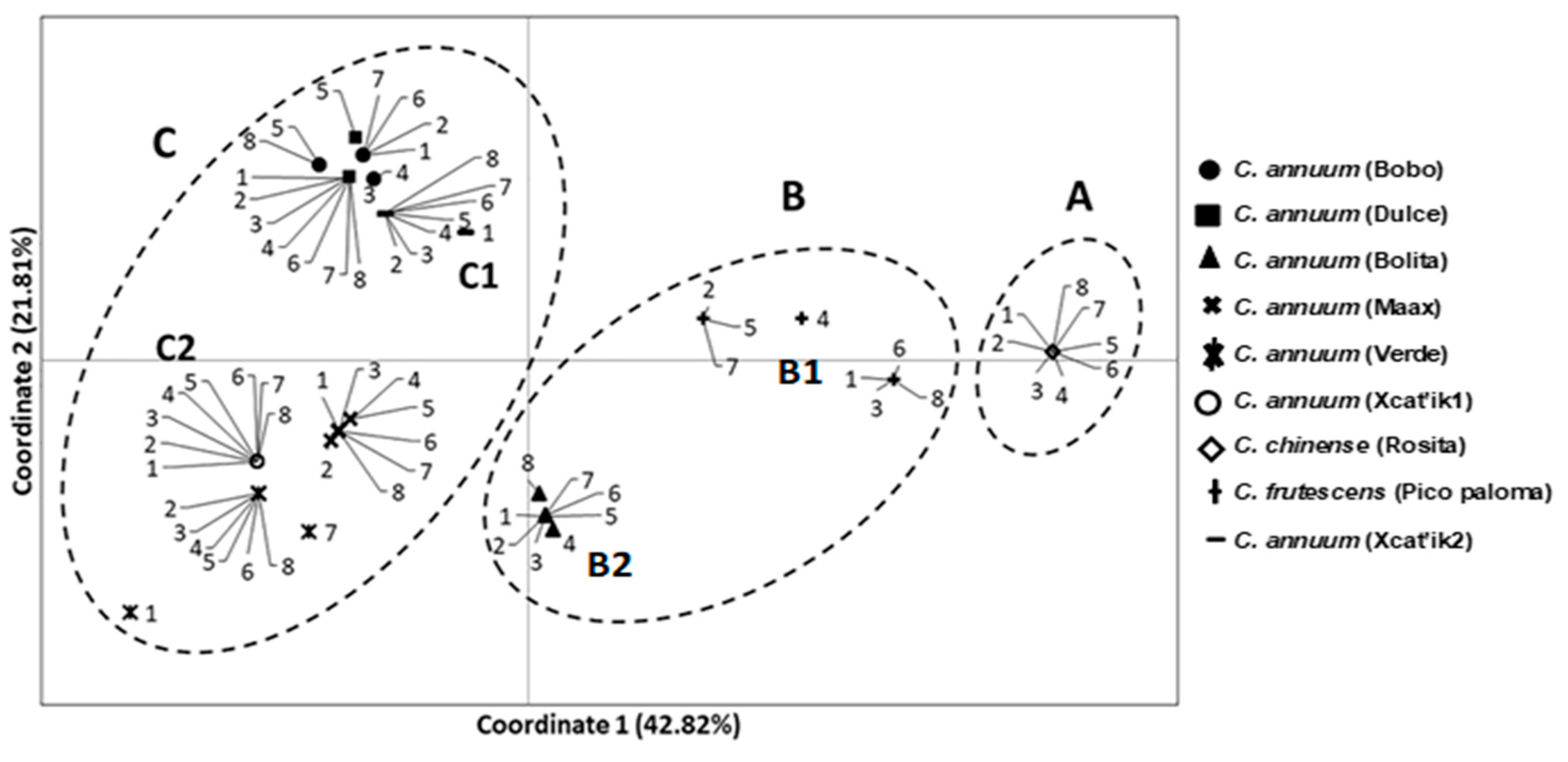
3.2. Genetic Relationships
3.3. Genetic Diversity
4. Discussion
4.1. Genetic Relationships between Nine Accessions of Capsicum Species Landraces from the Yucatan Peninsula
4.2. Genetic Diversity of Nine Accessions of Capsicum Species from Yucatan Peninsula
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- USDA-ARS. Grin Species Records of Capsicum; National Germplasm Resources Laboratory: Beltsville, MD, USA, 2011.
- Eshbaugh, W.H. Peppers: History and exploitation of a serendipitous new crop discovery. In New Crops; Janick, J., Simon, J.E., Eds.; Wiley: New York, NY, USA, 1993; pp. 132–139. [Google Scholar]
- Kraft, K.H.; Brown, C.H.; Nabhan, G.P.; Luedeling, E.; Luna Ruiz, J.J.; D’Eeckenbrugge, G.C.; Hijmans, R.J.; Gepts, P. Multiple lines of evidence for the origin of domesticated chili pepper, Capsicum annuum, in Mexico. Proc. Natl. Acad. Sci. USA 2014, 111, 6165–6170. [Google Scholar] [CrossRef]
- Rodriguéz, J.; Peña, O.B.V.; Gil, M.A.; Martínez, C.B.; Manzano, F.; Salazar, L.L. Rescate in situ de chile “Poblano” en Puebla, México. Rev. Fit. Mex. 2007, 30, 25–32. [Google Scholar]
- Hernández, V.S. Los parientes silvestres del chile: Su importancia biológica y cultural. In El chile: Protagonista de la independencia y la revolución; Richterich, K., Ed.; Fundación Herdez: México City, México, 2011; pp. 21–26. [Google Scholar]
- Aguilar-Meléndez, A.; Morrell, P.L.; Roose, M.L.; Kim, S.C. Genetic diversity and structure in semiwild and domesticated chiles (Capsicum annuum; Solanaceae) from Mexico. Am. J. Bot. 2009, 96, 1190–1202. [Google Scholar] [CrossRef] [PubMed]
- Latournerie, L.; Chávez, J.L.; Pérez, M.; Hernández, C.F.; Martínez, R.; Arias, L.M.; Castañón, G. Exploración de la diversidad morfológica de chiles regionales en Yaxcabá, Yucatán, México. Agron. Mesoam. 2001, 12, 41–47. [Google Scholar] [CrossRef]
- Ix-Nahuat, J.G.; Laournerie-Moreno, L.; Pech-May, A.M.; Pérez-Gutiérrez, A.; Tun-Suárez, J.M.; Ayora-Ricalde, G.; Mijangos-Cortes, J.O.; Castañón-Nájera, G.; López-Vázquez, J.S.; Montes-Hernández, S. Valor agronómico de germoplasma de chile dulce (Capsicum annuum L.) en Yucatán, México. Univ. Y Ciencia 2013, 29, 231–242. [Google Scholar]
- López-Espinosa, S.T.; Latournerie-Moreno, L.; Castañón-Nájera, G.; Ruiz-Sánchez, E.; Gómez-Leyva, J.F.; Andueza-Noh, R.H.; Mijangos-Cortés, J.O. Diversidad genética de chile habanero (Capsicum chinense jacq.) mediante ISSR. Rev. Fitotec. Mex. 2018, 41, 227–236. [Google Scholar]
- Laborde, C.J.A.; Pozo, O.C. Presente Y Pasado Del Chile En México; INIA-SARH: México City, México, 1982; p. 80. [Google Scholar]
- Rana, M.; Sharma, R.; Sharma, P.; Bhardwaj, S.V.; Sharma, M. Estimation of Genetic Diversity in Capsicum annuum L. Germplasm Using PCR-Based Molecular Markers. Natl. Acad. Sci. Lett. 2014, 37, 295–301. [Google Scholar] [CrossRef]
- Paterson, A.H.; Tanksley, S.D.; Sorrells, M.E. DNA markers in plant improvement. Adv. Agron. 1991, 46, 39–90. [Google Scholar] [CrossRef]
- Dias, G.B.; Gomes, V.M.; Moraes, T.M.S.; Zottich, U.P.; Rabelo, G.R.; Carvalho, A.O.; Moulin, M.L.S.; Goncalves, A.; Rodrigues, R.; Da Cunha, M. Characterization of Capsicum species using anatomical and molecular data. Genet. Mol. Res. 2013, 12, 6488–6501. [Google Scholar] [CrossRef]
- Carvalho, S.I.C.; Ragassi, C.F.; Bianchetti, L.B.; Reifschneider, F.J.B.; Buso, G.S.C.; Faleiro, F.G. Morphological and genetic relationships between wild and domesticated forms of peppers (Capsicum frutescens L. and C. chinense Jacquin). Genet. Mol. Res. 2014, 13, 7447–7464. [Google Scholar] [CrossRef]
- Martinez, A.L.A.; Araújo, J.S.P.; Ragassi, C.F.; Buso, G.S.C.; Reifschneider, F.J.B. Variability among Capsicum baccatum accessions from Góias, Brazil, assessed by morphological traits and molecular markers. Genet. Mol. Res. 2017, 16. [Google Scholar] [CrossRef] [PubMed]
- Zietkiewicz, E.; Rafalski, A.; Labuda, D. Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification. Genomics 1994, 20, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.; Doyle, J. A rapid procedure for DNA purification from small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Yao, H.; Zhao, Y.; Chen, D.F.; Chen, J.K.; Zhou, T.S. ISSR primer screening and preliminary evaluation of genetic diversity in wild populations of Glycyrrhiza Uralensis. Biol. Plant. 2008, 52, 117–120. [Google Scholar] [CrossRef]
- Thul, S.T.; Darokar, M.l.l.P.; Shasany, A.K.; Khanuja, A.K. Molecular Profiling for Genetic Variability in Capsicum Species Based on ISSR and RAPD Markers. Mol. Biotechnol. 2012, 51, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Earl, D.A.; Vonhold, M. Structure harvester: A web site and program for visualizing structure output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Palaeontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 4–9. [Google Scholar]
- Lynch, M.; Milligan, B.G. Analysis of population genetic structure with RAPD markers. Mol. Ecol. 1994, 3, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Krauss, S.L. Accurate gene diversity estimates from amplified fragment length polymorphism (AFLP) markers. Mol. Ecol. 2000, 9, 1241–1245. [Google Scholar] [CrossRef] [PubMed]
- Zhivotovsky, L. Estimating population structure in diploids with multilocus dominant DNA markers. Mol. Ecol. 1999, 8, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Yeh, F.C.; Boyle, T.J. POPGENE v. 1.31. Microsoft Windows-Based Freeware for Population Analysis; University of Alberta and Centre for International Forestry Research; University of Alberta: Edmonton, AB, Canada, 1999. [Google Scholar]
- Vekemans, X. AFLP-SURV Version 1.0. Distributed by the Author; Laboratoire de Génétique et Ecologie Végétale, Université Libre de Bruxelles: Brussels, Belgium, 2002. [Google Scholar]
- Hutchinson, J.B. Crop plant evolution: A general discussion. In Essays on Crop Plant Evolution; Hutchinson, J.B., Ed.; Cambridge University Press: Cambridge, UK, 1950; pp. 166–187. [Google Scholar]
- Jensen, R.J.; McLeod, M.J.; Eshbaugh, W.H.; Guttman, S.I. Numerical taxonomic analyses of allozymic variation in Capsicum (Solanaceae). Taxon 1979, 28, 315–327. [Google Scholar] [CrossRef]
- Albrecht, E.; Zhang, D.; Saftner, R.A.; Stommel, J.R. Genetic diversity and population structure of Capsicum baccatum genetic resources. Genet. Resour. Crop. Evol. 2012, 59, 517–538. [Google Scholar] [CrossRef]
- Hernández, V.S.; González, A.R.; Sánchez, P.P.; Casas, A.; Oyama, K. Estructura y diferenciación genética de poblaciones silvestres y domesticadas de chile del noroeste de México analizada con isoenzimas y RAPDS. Rev. Fitotec. Mex. 2006, 29, 25–29. [Google Scholar]
- Oyama, K.; Hernández-Verdugo, S.; Sánchez, C.; González-Rodríguez, A.; Sánchez-Peña, P.; Garzón-Tiznado, J.A.; Casas, A. Genetic structure of wild and domesticated populations of Capsicum annuum (Solanaceae) from northwestern Mexico analyzed by RAPDs. Genet. Resour. Crop. Evol. 2006, 53, 553–562. [Google Scholar] [CrossRef]
- Jarret, R.L. DNA Barcoding in a Crop Genebank: Resolving the Capsicum annuum species Complex. Open Biol. J. 2008, 1, 35–42. [Google Scholar] [CrossRef]
- Yumnam, J.S.; Tyagi, W.; Pandey, A.; Meetei, N.T.; Rai, M. Evaluation of genetic diversity of chilli landraces from North Eastern India based on morphology, SSR markers and the Pun1 locus. Plant. Mol. Biol. Rep. 2012, 30, 1470–1479. [Google Scholar] [CrossRef]
- Rai, V.P.; Kumar, R.; Kumar, S.; Rai, A.; Singh, M.; Singh, P.S.; Rai, A.B.; Paliwal, R. Genetic diversity in Capsicum germplasm based on microsatellite and random amplified microsatellite polymorphism markers. Physiol. Mol. Biol. Plants 2013, 19, 575–586. [Google Scholar] [CrossRef] [Green Version]
- Camacho-Pérez, L.; Martínez-Castillo, J.; Mijangos-Cortes, J.O.; Ferrer-Ortega, M.M.; Baudoin, J.P.; Andueza-Noh, R.H. Genetic structure of Lima bean (Phaseolus lunatus L.) landraces grown in the Mayan area. Genet. Resour. Crop. Evol. 2018, 65, 229–241. [Google Scholar] [CrossRef]
- Martínez-Castillo, J.; Colunga-García Marín, P.; Zizumbo-Villarreal, D. Genetic erosion and in situ conservation of Lima bean (Phaseolus lunatus L.) landraces in its Mesoamerican diversity center. Genet. Resour. Crop. Evol. 2008, 55, 1065–1077. [Google Scholar] [CrossRef]
- González, E.T.; Casanova, C.C.; Gutiérrez, L.P.; Torres, L.T.; Contreras, F.M.; Peraza, S.S. Chiles cultivados en Yucatán. In Biodiversidad y Desarrollo.Humano en Yucatán; Durán, R., Méndez, M., Eds.; CICY CONABIO, SEDUMA: Yucatán, Mexico, 2010; pp. 342–344. [Google Scholar]
- Baral, J.B.; Bosland, P.W. Unraveling the species dilemma in Capsicum frutescens and C. chinense (Solanaceae): A multiple evidence approach using morphology, molecular analysis, and sexual compatibility. J. Amer. Soc. Hort. Sci. 2004, 129, 826–832. [Google Scholar] [CrossRef]
- Ince, A.G.; Karaca, M.; Onus, A.N. Genetic relationships within and between Capsicum species. Biochem. Genet. 2010, 48, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Soria-Fregoso, M.; Trejo-Rivero, J.A.; Tun-Suárez, J.M.; Terán-Saldivar, R. Paquete TecnolóGico Para la Producción de Chile Habanero, 3rd ed.; SEP, DGETA ITA-2: Yucatán, México, 2002; p. 430. [Google Scholar]
- Pickersgill, B. The archeological record of chili peppers (Capsicum spp.) and the secuence of the domestication in Peru. Amer. Antiq. 1969, 34, 54–61. [Google Scholar] [CrossRef]
- Tanksley, S.D.; McCouch, S.R. Seed banks and molecular maps: Unlocking genetic potential from the wild. Science 1997, 277, 1063–1066. [Google Scholar] [CrossRef]
- Hanáček, P.; Vyhnánek, T.; Rohrer, M.; Cieslarová, J.; Stavĕlíková, H. DNA polymorphism in genetic resources of red pepper using microsatellite markers. Hort. Sci. 2009, 36, 127–132. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Df | SS | Variance Component | Variation Percentage |
|---|---|---|---|---|
| Among accessions | 8 | 416.972 | 6.449 | 92 |
| Within accessions | 63 | 33.375 | 0.530 | 8 |
| Total | 71 | 450.347 | 6.979 | 100 |
| Accession | %P | Na | Ne | I | HBay |
|---|---|---|---|---|---|
| Bobo | 4.76 | 1.0476 | 1.0476 | 0.0330 | 0.02009 |
| Dulce | 7.14 | 1.0714 | 1.0601 | 0.0464 | 0.01758 |
| Maax | 4.76 | 1.0476 | 1.0271 | 0.0222 | 0.01590 |
| Verde | 2.38 | 1.0238 | 1.0211 | 0.0158 | 0.00666 |
| Xcat’ik1 | 0.00 | 1.000 | 1.000 | 0.000 | 0.00000 |
| Xcat’ik2 | 4.76 | 1.0476 | 1.0401 | 0.0309 | 0.01172 |
| Bolita | 4.76 | 1.0476 | 1.0401 | 0.0309 | 0.01172 |
| Pico Paloma | 14.29 | 1.1429 | 1.1100 | 0.0887 | 0.07701 |
| Rosita | 0.00 | 1.000 | 1.000 | 0.000 | 0.00000 |
| Total | 88.10 | 1.8810 | 1.5107 | 0.4402 | 0.0176 |
| Group | No. Samples | %P | Na | Ne | I | HBay |
|---|---|---|---|---|---|---|
| 1 | 48 | 54.76 | 1.5476 | 1.3614 | 0.2935 | 0.19991 |
| 2 | 16 | 38.10 | 1.3810 | 1.3078 | 0.2376 | 0.17590 |
| 3 | 8 | 0.00 | 1.000 | 1.000 | 0.000 | 0.00000 |
| Total | 72 | 88.10 | 1.8810 | 1.5107 | 0.4402 | 0.0179 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
López Castilla, L.d.C.; Garruña Hernández, R.; Castillo Aguilar, C.d.l.C.; Martínez-Hernández, A.; Ortiz-García, M.M.; Andueza-Noh, R.H. Structure and Genetic Diversity of Nine Important Landraces of Capsicum Species Cultivated in the Yucatan Peninsula, Mexico. Agronomy 2019, 9, 376. https://doi.org/10.3390/agronomy9070376
López Castilla LdC, Garruña Hernández R, Castillo Aguilar CdlC, Martínez-Hernández A, Ortiz-García MM, Andueza-Noh RH. Structure and Genetic Diversity of Nine Important Landraces of Capsicum Species Cultivated in the Yucatan Peninsula, Mexico. Agronomy. 2019; 9(7):376. https://doi.org/10.3390/agronomy9070376
Chicago/Turabian StyleLópez Castilla, Lucero del C., René Garruña Hernández, Crescencio de la Cruz Castillo Aguilar, Aida Martínez-Hernández, Matilde Margarita Ortiz-García, and Rubén H. Andueza-Noh. 2019. "Structure and Genetic Diversity of Nine Important Landraces of Capsicum Species Cultivated in the Yucatan Peninsula, Mexico" Agronomy 9, no. 7: 376. https://doi.org/10.3390/agronomy9070376