Bacterial Communities in the Rhizosphere and Phyllosphere of Halophytes and Drought-Tolerant Plants in Mediterranean Ecosystems

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Sites and Sample Collection
2.2. Soil Properties
2.3. Sample Processing and Sequencing
2.4. Data Analysis
2.5. Read Processing
3. Results
3.1. Soil Properties in the Sampling Sites
3.2. Bacterial Communities’ Diversity and Composition
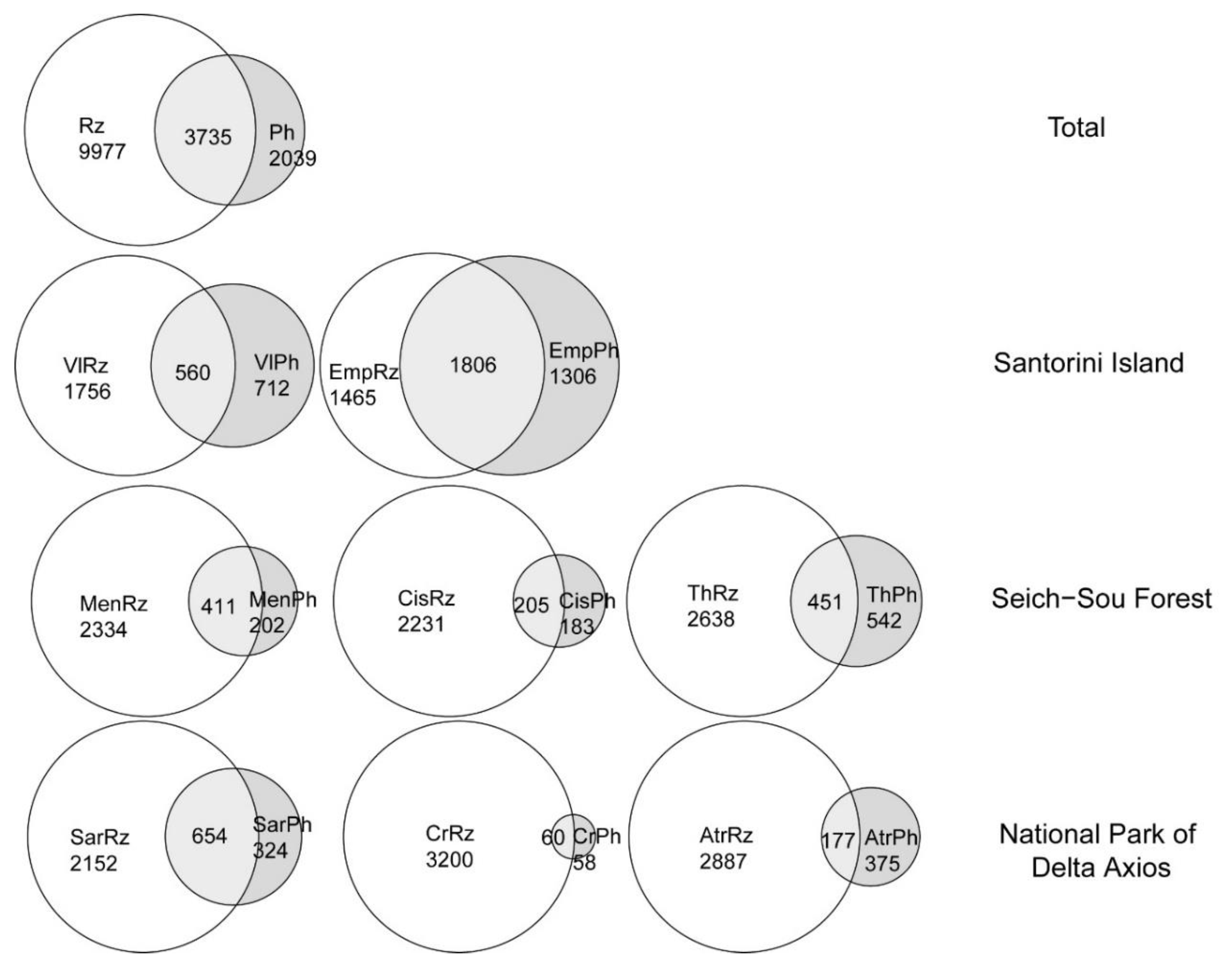
3.3. Rhizosphere vs. Phyllosphere and Spatial Heterogeneity
3.4. Generalists and Abundant Specialists
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Buée, M.; De Boer, W.; Martin, F.; Van Overbeek, L.; Jurkevitch, E. The rhizosphere zoo: An overview of plant-associated communities of microorganisms, including phages, bacteria, archaea, and fungi, and some of their structuring factors. Plant Soil 2009, 321, 189–212. [Google Scholar] [CrossRef]
- Kent, A.D.; Triplett, E.W. Microbial communities and their interactions in soil and rhizosphere ecosystems. Annu. Rev. Microbiol. 2002, 56, 211–236. [Google Scholar] [CrossRef] [PubMed]
- Kai, M.; Effmert, U.; Piechulla, B. Bacterial-plant-interactions: Approaches to unravel the biological function of bacterial volatiles in the rhizosphere. Front. Microbiol. 2016, 7, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kai, M.; Piechulla, B. Plant growth promotion due to rhizobacterial volatiles—An effect of CO2? FEBS Lett. 2009, 583, 3473–3477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Bringel, F.; Couée, I. Pivotal roles of phyllosphere microorganisms at the interface between plant functioning and atmospheric trace gas dynamics. Front. Microbiol. 2015, 6, 486. [Google Scholar] [CrossRef] [Green Version]
- Brussaard, L. Biodiversity and ecosystem functioning in soil. Ambio 1997, 26, 563–570. [Google Scholar]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Forest soil bacteria: Diversity, involvement in ecosystem processes, and response to global change. Microbiol. Mol. Biol. Rev. 2017, 81, e00063-16. [Google Scholar] [CrossRef] [Green Version]
- Bruto, M.; Prigent-Combaret, C.; Muller, D.; Moënne-Loccoz, Y. Analysis of genes contributing to plant-beneficial functions in plant growth-promoting rhizobacteria and related Proteobacteria. Sci. Rep. 2014, 4, 6261. [Google Scholar] [CrossRef] [Green Version]
- Rajput, L.; Imran, A.; Mubeen, F.; Hafeez, F.Y. Salt-tolerant PGPR strain Planococcus rifietoensis promotes the growth and yield of wheat (Triticum aestivum L.) cultivated in saline soil. Pak. J. Bot. 2013, 45, 1955–1962. [Google Scholar]
- Sharma, S.; Kulkarni, J.; Jha, B. Halotolerant rhizobacteria promote growth and enhance salinity tolerance in peanut. Front. Microbiol. 2016, 7, 1600. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.P.; Jha, P.N. The multifarious PGPR Serratia marcescens CD-13 augments induced systemic resistance and enhanced salinity tolerance of wheat (Triticum aestivum L.). PLoS ONE 2016, 11, e0155026. [Google Scholar] [CrossRef]
- Yadav, V.K.; Raghav, M.; Sharma, S.K.; Bhagat, N. Rhizobacteriome: Promising candidate for conferring drought tolerance in crops. J. Pure Appl. Microbiol. 2020, 14, 73–92. [Google Scholar] [CrossRef] [Green Version]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of action of plant growth promoting bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef] [Green Version]
- Mukhtar, S.; Malik, K.A.; Mehnaz, S. Microbiome of Halophytes: Diversity and importance for plant health and productivity. Microbiol. Biotechnol. Lett. 2019, 47, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Andreote, F.D.; e Silva, M.C.P. Microbial communities associated with plants: Learning from nature to apply it in agriculture. Curr. Opin. Microbiol. 2017, 37, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Schlemper, T.R.; Leite, M.F.A.; Lucheta, A.R.; Shimels, M.; Bouwmeester, H.J.; Van Veen, J.A.; Kuramae, E.E. Rhizobacterial community structure differences among sorghum cultivars in different growth stages and soils. FEMS Microbiol. Ecol. 2017, 93, 96. [Google Scholar] [CrossRef] [Green Version]
- Vacher, C.; Hampe, A.; Porté, A.J.; Sauer, U.; Company, S.; Morris, C.E. The phyllosphere: Microbial jungle at the plant-climate interface. Annu. Rev. Ecol. Evol. S. 2016, 47, 1–24. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Van Themaat, E.V.L.; Schulze-Lefert, P. Structure and functions on the bacterial microbiota of plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, G.; Coaker, G.L.; Leveau, J.H.J. New insights into the structure and function of phyllosphere microbiota through high-throughput molecular approaches. FEMS Microbiol. Lett. 2013, 348, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barret, M.; Briand, M.; Bonneau, S.; Préveaux, A.; Valière, S.; Bouchez, O.; Hunault, G.; Simoneau, P.; Jacquesa, M.-A. Emergence shapes the structure of the seed microbiota. Appl. Environ. Microb. 2015, 81, 1257–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassani, M.A.; Durán, P.; Hacquard, S. Microbial interactions withing the plant holobiont. Microbiome 2018, 6, 58. [Google Scholar] [CrossRef]
- Garza, D.R.; Dutilh, B.E. From cultured to uncultured genome sequences: Metagenomes and modelling microbial ecosystems. Cell. Mol. Life Sci. 2015, 72, 4287–4308. [Google Scholar] [CrossRef] [Green Version]
- Bálint, M.; Bahram, M.; Eren, A.M.; Faust, K.; Fuhrman, J.A.; Lindahl, B.; O’Hara, R.B.; Öpik, M.; Sogin, M.L. Millions of reads, thousands of taxa: Microbial community structure and associations analyzed via marker genesa. FEMS Microbiol. Rev. 2016, 40, 686–700. [Google Scholar] [CrossRef] [Green Version]
- Mauchline, T.H.; Malone, J.G. Life in earth—The root microbiome to the rescue? Curr. Opin. Microbiol. 2017, 37, 23–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, I.; Ventorino, V.; Pepe, O. Effectiveness of plant beneficial microbes: Overview of the methodological approaches for the assessment of root colonization and persistence. Front. Plant Sci. 2020, 11, 6. [Google Scholar] [CrossRef] [Green Version]
- White, R.A.; Rivas-Ubach, A.; Borkum, M.I.; Köberl, M.; Bilbao, A.; Colby, S.M.; Hoyt, D.W.; Bingol, K.; Kim, Y.-M.; Wendler, J.P.; et al. The state of rhizospheric science in the era of multi-omics: A practical guide to omics technologies. Rhizosphere 2017, 3, 212–221. [Google Scholar] [CrossRef]
- Beilsmith, K.; Thoen, M.P.M.; Brachi, B.; Gloss, A.D.; Khan, M.H.; Begelson, J. Genome-wide association studies on the phyllosphere microbiome: Embracing complexity in host-microbe interactions. Plant J. 2019, 97, 164–181. [Google Scholar] [CrossRef]
- Vokou, D.; Giannakou, U.; Kontaxi, C.; Vareltzidou, S. Aliakmon and Gallikos Delta Complex, Northern Greece. In Encyclopedia of Wetlands; Finlayson, M., Prentice, C., Milton, R., Eds.; Springer: Geneva, Switzerland, 2016; Volume 4, ISBN 13: 9789400740006. [Google Scholar]
- Bouyoucos, G.J. Hydrometer method improved for making particle size analysis of soils. Agron. J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- Sparks, D.L. Methods of Soil Analysis—Part 3—Chemical Methods; SSSA Book Series 5: Madison, WI, USA, 1996. [Google Scholar]
- Mellidou, I.; Moschou, P.N.; Ioannidis, N.E.; Pankou, C.; Gèmes, K.; Valassakis, C.; Andronis, E.A.; Beris, D.; Haralampidis, K.; Roussis, A.; et al. Silencing S-adenosyl-L-methionine decarboxylase (SAMDC) in Nicotiana tabacum points at a polyamine-dependent trade-off between growth and tolerance responses. Front. Plant Sci. 2016, 7, 379. [Google Scholar] [CrossRef] [Green Version]
- Allison, L.E.; Moodie, C.D. Volumetric calsimeter methods of soil analysis, part 2. Chemical and microbiological properties. In American Society of Agronomy; Black, C.A., Ed.; Information Age Publishing: Charlotte, NC, USA, 1965. [Google Scholar]
- Mulvaney, P. Surface plasmon spectroscopy of nanosized metal particles. Langmuir 1996, 12, 788–800. [Google Scholar] [CrossRef]
- Kuo, S. Phosphorus. In Methods in Soil Analysis, Part 3: Chemical Methods; Sparks, D.L., Ed.; SSSA: Madison, WI, USA, 1996. [Google Scholar]
- Thomas, G.W. Exchangeable cations. In Methods of Soil Analysis, Part 2; Page, A.L., Ed.; Agronomy Monographs: Madison, WI, USA, 1982; Volume 9, pp. 159–165. [Google Scholar]
- Comeau, A.M.; Li, W.K.; Tremblay, J.E.; Carmack, E.C.; Lovejoy, C. Arctic Ocean microbial community structure before and after the 2007 record sea ice minimum. PLoS ONE 2011, 6, e27492. [Google Scholar] [CrossRef] [PubMed]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Paleontol. Electron. 2001, 4, 9. [Google Scholar]
- Baselga, A.; Orme, C.D.L. Betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Koleff, P.; Gaston, K.J.; Lennon, J.J. Measuring beta diversity for presence-absence data. J. Anim. Ecol. 2003, 72, 367–382. [Google Scholar] [CrossRef] [Green Version]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Primer v6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- Levins, R. Evolution in Changing Environments; Princeton University Press: Princeton, NJ, USA, 1968. [Google Scholar]
- Pandit, S.N.; Kolasa, J.; Cottenie, K. Contrasts between habitat generalists and specialists: An empirical extension to the basic metacommunity framework. Ecology 2009, 90, 2253–2562. [Google Scholar] [CrossRef] [Green Version]
- Székely, A.J.; Langenheder, S. The importance of species sorting differs between habitat generalists and specialists in bacterial communities. FEMS Microbiol. Ecol. 2014, 87, 102–112. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microb. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Gevers, D.; Westcott, S.L. Reducing the effects of PCR amplification and sequencing artifacts on 16S rRNA-based studies. PLoS ONE 2011, 6, e27310. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, 590–596. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Kunin, V.; Engelbrektson, A.; Ochman, H.; Hugenholtz, P. Wrinkles in the rare biosphere: Pyrosequencing errors can lead to artificial inflation of diversity estimates. Environ. Microbiol. 2010, 12, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Richards, T.A.; Leonard, G.; Mahé, F.; Del Campo, J.; Romac, S.; Jones, M.D.M.; Maguire, F.; Dunthorn, M.; De Vargas, C.; Massana, R.; et al. Molecular diversity and distribution of marine fungi across 130 European environmental samples. Proc. R. Soc. B 2015, 282, 20152243. [Google Scholar] [CrossRef]
- Leontidou, K.; Genitsaris, S.; Papadopoulou, A.; Kamou, N.; Bosmali, I.; Matsi, T.; Madesis, P.; Vokou, D.; Karamanoli, K.; Mellidou, I.; et al. Plant growth promoting rhizobacteria isolated from halophytes and drought-tolerant plants: Genomic characterization and exploration of phyto-beneficial traits. Sci. Rep. 2020, 10, 14857. [Google Scholar] [CrossRef] [PubMed]
- Knief, C.; Delmotte, N.; Chaffron, S.; Stark, M.; Innerebner, G.; Wassmann, R.; Von Mering, C.; Vorholt, J.A. Metaproteogenomic analysis of microbial communities in the phyllosphere and rhizosphere of rice. ISME J. 2012, 6, 1378–1390. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Singh, D.; Lai-Hoe, A.; Go, R.; Rahim, R.A.; Ainuddin, A.N.; Chun, J.; Adams, J.M. Distinctive phyllosphere bacterial communities in tropical trees. Microb. Ecol. 2012, 63, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Lambais, M.R.; Lucheta, A.R.; Crowley, D.E. Bacterial community assemblages associated with the phyllosphere, dermosphere, and rhizosphere of tree species of the Atlantic Forest are host taxon dependent. Microb. Ecol. 2014, 68, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.R.; Lundberg, D.S.; Del Rio, T.G.; Tringe, S.G.; Dangl, J.L.; Mitchell-Olds, T. Host genotype and age shape the leaf and root microbiomes of a wild perennial plant. Nat. Commun. 2016, 7, 12151. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.; Laforest-Lapointe, I.; Kembel, S.W. Variation in the leaf and root microbiome of sugar maple (Acer saccharum) at an elevational range limit. PeerJ 2018, e5293. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.J.; Wang, L.L.; Li, Q.; Shang, Q.M. Bacterial communities in the rhizosphere, phyllosphere and endosphere of tomato plants. PLoS ONE 2019, 14, e0223847. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Zhang, X.; He, R.; Wang, S.; Jiao, C.; Huang, R.; He, X.; Zeng, J.; Zhao, D. The composition and assembly of bacterial communities across the rhizosphere and phyllosphere compartments of Phragmites australis. Diversity 2019, 11, 98. [Google Scholar] [CrossRef] [Green Version]
- Bao, L.; Cai, W.; Cao, J.; Zhang, X.; Liu, J.; Chen, H.; Wei, Y.; Zhuang, X.; Zhuang, G.; Bai, Z.; et al. Microbial community overlap between the phyllosphere and rhizosphere of three plants from Yongxing Island, South China Sea. Microbiol. Open 2020, 9, e1048. [Google Scholar] [CrossRef] [PubMed]
- Venturi, V.; Keel, C. Signaling in the rhizosphere. Trends Plant Sci. 2016, 21, 187–198. [Google Scholar] [CrossRef]
- Beattie, G.A.; Lindow, S.E. Bacterial colonization of leaves: A spectrum of strategies. Phytopathology 1999, 89, 353–359. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [Green Version]
- Lindström, E.S.; Langenheder, S. Local and regional factors influencing bacterial community assembly. Environ. Microbiol. Rep. 2012, 4, 1–9. [Google Scholar] [CrossRef]
- Nemergut, D.R.; Costello, E.K.; Hamady, M.; Lozupone, C.; Jiang, L.; Schmidt, S.K.; Townsend, A.R.; Cleveland, C.C.; Stanish, L.; Knight, R.; et al. Global patterns in the biogeography of bacterial taxa. Environ. Microbiol. 2011, 13, 135–144. [Google Scholar] [CrossRef]
- Economou, A.; Skouteri, A.; Michopoulos, P. Soils and land use of Santorini, Greece. In Soils of Volcanic Regions in Europe; Arnalds, Ó., Óskarsson, H., Bartoli, F., Buurman, P., Stoops, G., García-Rodeja, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Lindow, S.E.; Brandl, M.T. Microbiology of the phyllosphere. Appl. Environ. Microb. 2003, 69, 1875–1883. [Google Scholar] [CrossRef] [Green Version]
- Mello, B.; Alessi, A.M.; McQueen-Mason, S.; Bruce, N.C.; Polikarpov, I. Nutrient availability shapes the microbial community structure in sugarcane bagasse compost-derived consortia. Sci. Rep. 2016, 6, e38781. [Google Scholar] [CrossRef] [Green Version]
- Álvarez-López, V.; Prieto-Fernández, Á.; Becerra-Castro, C.; Monterroso, C.; Kidd, P.S. Rhizobacterial communities associated with the flora of three serpentine outcrops of the Iberian Peninsula. Plant Soil 2016, 403, 233–252. [Google Scholar] [CrossRef]
- Jorquera, M.A.; Maruyama, F.; Ogram, A.V.; Navarette, O.U.; Lagos, L.M.; Inostroza, N.G.; Acuña, J.J.; Rilling, J.I.; De La Luz Mora, M. Rhizobacterial community structures associated with native plants grown in Chilean extreme environments. Microb. Ecol. 2016, 72, 633–646. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.N.; Verma, P.; Kumar, S.; Kumar, V.; Kumar, M.; Kumari Sugitha, T.C.; Singh, B.P.; Saxena, A.K.; Dhaliwal, H.S. Actinobacteria from rhizosphere: Molecular diversity, distributions, and potential biotechnological applications. In New and Future Developments in Microbial Biotechnology and Bioengineering: Actinobacteria: Diversity and Biotechnological Applications; Singh, B.P., Gupta, V., Passari, A.K., Eds.; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Vokou, D.; Genitsaris, S.; Karamanoli, K.; Vareli, K.; Zachari, M.; Voggoli, D.; Monokrousos, N.; Halley, J.M.; Sainis, I. Metagenomic characterization reveals pronounced seasonality in the diversity and structure of the phyllosphere bacterial community in a Mediterranean ecosystem. Microorganisms 2019, 7, 518. [Google Scholar] [CrossRef] [Green Version]
- Han, H.S.; Lee, K.D. Plant growth promoting rhizobacteria effect on antioxidant status, photosynthesis, mineral uptake and growth of lettuce under soil salinity. Res. J. Agric. Biol. Sci. 2005, 1, 210–215. [Google Scholar]
- Sakai, M.; Hosoda, A.; Ogura, K.; Ikenaga, M. The growth of Steroidobacter agariperforans sp. nov., a novel agar-degrading bacterium isolated from soil, is enhanced by the diffusible metabolites produced by bacteria belonging to rhizobiales. Microbes Environ. 2014, 29, 89–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velázquez-Becerra, C.; Macías-Rodríguez, L.I.; López-Bucio, J.; Altamirano-Hernández, J.; Flores-Cortez, I.; Valencia-Cantero, E. A volatile organic compound analysis from Arthrobacter agilis identifies dimethylhexadecylamine, an amino-containing lipid modulating bacterial growth and Medicago sativa morphogenesis in vitro. Plant Soil 2011, 339, 329–340. [Google Scholar] [CrossRef]
- Sashidhar, B.; Podile, A.R. Mineral phosphate solubilization by rhizosphere bacteria and scope for manipulation of the direct oxidation pathway involving glucose dehydrogenase. J. Appl. Microbiol. 2010, 109, 1–12. [Google Scholar] [CrossRef]
- Emmanuel, E.S.C.; Ananthi, T.; Anandkumar, B.; Maruthamuthu, S. Accumulation of rare earth elements by siderophore-forming Arthrobacter luteolus isolated from rare earth environment of Chavara, India. J. Biosci. 2012, 37, 25–31. [Google Scholar] [CrossRef]
- Requena, N.; Jimenez, I.; Toro, M.; Barea, J.M. Interactions between plant-growth-promoting rhizobacteria (PGPR), arbuscular mycorrhizal fungi and Rhizobium spp. in the rhizosphere of Anthyllis cytisoides, a model legume for revegetation in Mediterranean semi-arid ecosystems. New Phytol. 1997, 136, 667–677. [Google Scholar] [CrossRef]
- Timmusk, S.; Nicander, B.; Granhall, U.; Tillberg, E. Cytokinin production by Paenibacillus polymyxa. Soil Biol. Biochem. 1999, 31, 1847–1852. [Google Scholar] [CrossRef]
- Townsend, C.; Begon, M.; Harper, J. Essentials of Ecology, 2nd ed.; Blackwell Publishing: Oxford, UK, 2003. [Google Scholar]
- Van der Gucht, K.; Cottenie, K.; Muylaert, K.; Vloemans, N.; Cousin, S.; Declerck, S.; Jeppesen, E.; Conde-Porcuna, J.M.; Schwenk, K.; Zwart, G.; et al. The power of species sorting: Local factors drive bacterial community composition over a wide range of spatial scales. Proc. Natl. Acad. Sci. USA 2007, 104, 20404–20409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Liu, J.J.; Banerjee, S.; Zhou, N.; Zhao, Z.Y.; Zhang, K.; Tian, C.Y. Soil pH is equally important as salinity in shaping bacterial communities in saline soils under halophytic vegetation. Sci. Rep. 2018, 8, 4550. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Wang, Y.; Zhang, L.; Tang, Y.; Luo, X.; An, H.; Fang, C. Hymenobacter tibetensis sp. nov., a UV-resistant bacterium isolated from Qinghai-Tiber plateau. Syst. Appl. Microbiol. 2009, 32, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Jorge, G.L.; Kisiala, A.; Morrison, E.; Aoki, M.; Nogueira, A.P.O.; Emery, R.J.N. Endosymbiotic Methylobacterium oryzae mitigates the impact of limited water availability in lentil (Lens culinaris Medik.) by increasing plant cytokinin levels. Environ. Exp. Bot. 2019, 162, 525–540. [Google Scholar] [CrossRef]
- Cohen, A.C.; Piccoli, P.; Bottini, R.; Salomon, M.V. Plant-rhizobacteria interaction and drought stress tolerance in plants. In Drought Stress Tolerance in Plants; Hossain, M., Wani, S., Bhattacharjee, S., Burritt, D., Tran, L.S., Eds.; Springer: Cham, Switzerland, 2016; Volume 1. [Google Scholar]
- Donhauser, J.; Niklaus, P.A.; Rousk, J.; Larose, C.; Frey, B. Temperatures beyond the community optimum promote the dominance of heat-adapted, fast growing and stress resistant bacteria in alpine soils. Soil Biol. Biochem. 2020, 148, 107873. [Google Scholar] [CrossRef]
- Choi, D.H.; Yi, H.; Chun, J.; Cho, B.C. Jannaschia seosinensis sp. nov., isolated from hypersaline water of a solar saltern in Korea. Int. J. Syst. Evol. Micr. 2006, 56, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Nakano, M.; Inagaki, T.; Okunishi, S.; Tanaka, R.; Maeda, H. Effect of salinity on denitrification under limited single carbon source by Marinobacter sp. isolated from marine sediment. J. Basic Microb. 2010, 50, 285–289. [Google Scholar] [CrossRef]
- Kang, H.; Kim, H.; Joung, Y.; Joh, K. Lewinella maritima sp. nov., and Lewinella lacunae sp. nov., novel bacteria from marine environments. Int. J. Syst. Evol. Micr. 2017, 67, 3603–3609. [Google Scholar] [CrossRef]
- Lu, D.C.; Xia, J.; Dunlap, C.A.; Rooney, A.P.; Du, Z.J. Gracilimonas halophila sp. nov., isolated from a marine solar saltern. Int. J. Syst. Evol. Micr. 2017, 67, 3251–3255. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Sites | Latitude (N) | Longitude (E) | Plant Species | Plant Compartment | Number of Samples Sequenced/Analysed after Denoizing | Code |
|---|---|---|---|---|---|---|
| Santorini—Vlichada Santorini—Emporio | 36°20′42″ | 25°26′44″ | Solanum lycopersicum, ‘Santorini’ (landrace) | Rhizosphere | 8/8 | VlRz |
| Phyllosphere | 8/8 | VlPh | ||||
| 36°20′57″ | 25°25′51″ | Rhizosphere | 8/8 | EmpRz | ||
| Phyllosphere | 8/8 | EmpPh | ||||
| Seich-Sou Forest | 40°37′41″ | 22°58′15″ | Mentha pulegium | Rhizosphere | 5/5 | MenRz |
| Phyllosphere | 5/4 | MenPh | ||||
| Cistus sp. | Rhizosphere | 5/4 | CisRz | |||
| Phyllosphere | 5/1 | CisPh | ||||
| Thymus sp. | Rhizosphere | 5/4 | ThRz | |||
| Phyllosphere | 5/4 | ThPh | ||||
| National Park of Delta Axios | 40°31′19″ | 22°39′02″ | Sarcocornia sp. | Rhizosphere | 5/5 | SarRz |
| Phyllosphere | 5/4 | SarPh | ||||
| Crithmum sp. | Rhizosphere | 5/5 | CrRZ | |||
| Phyllosphere | 5/2 | CrPh | ||||
| Atriplex sp. | Rhizosphere | 5/5 | AtrRz | |||
| Phyllosphere | 5/2 | AtrPh |
| BSOR | BSIM | BNES | |
|---|---|---|---|
| Santorini Island | 0.87/0.91 | 0.81/0.82 | 0.06/0.09 |
| Seich-Sou Forest | 0.88/0.85 | 0.84/0.77 | 0.04/0.08 |
| National Park of Delta Axios | 0.89/0.89 | 0.87/0.81 | 0.02/0.08 |
| OTUs | Bacterial Phylum or Class | Closest Relative (% Similarity) [Accession Number] | Isolation Source |
|---|---|---|---|
| OTU_11 | Actinobacteria | Blastococcus sp. (99.5%) [MK239642] | Granite building |
| OTU_14 | Alphaproteobacteria | Uncultured Sphingomonadaceae (98.9%) [KC329595] | Ginger cropping soil |
| OTU_20 | Alphaproteobacteria | Sphingomonas yunnanensis (99.3%) [MN968938] | Culture strain |
| OTU_23 | Alphaproteobacteria | Bradyrhizobium jicamae (99.3%) [KJ831347] | Culture strain |
| OTU_27 | Gammaproteobacteria | Steroidobacter sp. (99.1%) [MK311353] | Farmland soil |
| OTU_32 | Unidentified | Uncultured bacterium (99.3%) [KU191639] | Bryophyte |
| OTU_34 | Actinobacteria | Arthrobacter sp. (99.8%) [MK212372] | Soil |
| OTU_36 | Actinobacteria | Actinoplanes luteus (99.5%) [NR_145623] | Soil |
| OTU_39 | Alphaproteobacteria | Skermanella aerolata (98.6%) [MH259920] | Sargassum horneri |
| OTU_40 | Actinobacteria | Mycobacterium sp. (99.1%) [JX273679] | Root |
| OTU_44 | Actinobacteria | Solirubrobacter phytolaccae (98.6%) [MN686629] | Root |
| OTU_45 | Actinobacteria | Kribbella sandramycini (99.1%) [MT072122] | Soil |
| OTU_55 | Actinobacteria | Mycobacterium sp. (99.5%) [KX900598] | Culture strain |
| OTU_65 | Actinobacteria | Uncultured Rubrobacter sp. (99.5%) [KC110942] | Soil |
| OTU_69 | Actinobacteria | Pseudonocardia sp. (98.6%) [MN493045] | Root |
| OTU_72 | Unidentified | Uncultured bacterium (99.3%) [JN178597] | Extreme saline-alkaline soil |
| OTU_78 | Unidentified | Uncultured bacterium (98.4%) [JQ978633] | Permafrost soil |
| OTU_83 | Bacteroidetes | Uncultured Chitinophagaceae sp. (98.6%) [LN680465] | Coalmine overburden |
| OTU_86 | Unidentified | Uncultured bacterium (98.4%) [MH445072] | Rhizospheric soil |
| OTU_87 | Actinobacteria | Geodermatophilus sp. (97.9%) [MG200148] | Marine sponges |
| OTU_91 | Unidentified | Uncultured bacterium (99.5%) [MN175141] | Soil |
| OTU_120 | Alphaproteobacteria | Rhizobium sp. (99.8%) [MT023038] | Culture strain |
| OTU_122 | Alphaproteobacteria | Bosea sp. (98.2%) [AJ968693] | Culture strain |
| OTU_195 | Unidentified | Uncultured bacterium (97.9%) [EU172577] | Air |
| OTU_206 | Unidentified | Uncultured bacterium (99.1%) [KR560009] | Soil |
| OTU_212 | Unidentified | Uncultured bacterium (98.4%) [KP280904] | Root |
| OTU_217 | Alphaproteobacteria | Neorhizobium alkalisoli (99.8%) [KF580864] | Root |
| OTU_231 | Actinobacteria | Geodermatophilus aquaeductus (98.9%) [NR_136840] | Stone |
| OTU_234 | Unidentified | Uncultured bacterium (99.5%) [JF914288] | Seed |
| OTU_332 | Unidentified | Uncultured bacterium (99.1%) [KC331318] | Apple orchard |
| OTU_370 | Actinobacteria | Streptomyces sp. (99.1%) [MK638452] | Rhizosphere |
| OTU_372 | Unidentified | Uncultured bacterium (98.6%) [JQ049231] | Soil |
| OTU_380 | Unidentified | Uncultured bacterium (99.5%) [MF113653] | Dairy pasteurizer |
| OTU_412 | Firmicutes | Paenibacillus sp. (99.8%) [KC404044] | Wood core |
| OTU_461 | Alphaproteobacteria | Sphingomonas sp. (99.3%) [AJ968701] | Culture strain |
| OTU_709 | Unidentified | Uncultured bacterium (98.9%) [AB473917] | Endolithic system |
| Santorini Island | Seich-Sou Forest | National Park of Delta Axios | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OTUs | Bacterial Phylum or Class | Closest Relative (% Similarity) [Accession Number] | Isolation Source | Vl Rz | Vl Ph | EmpRz | EmpPh | MenRz | Men Ph | Cis Rz | Cis Ph | Th Rz | Th Ph | Sar Rz | Sar Ph | Cr Rz | Cr Ph | Atr Rz | Atr Ph |
| OTU_2 | Alphaproteobacteria | Sphingobium sp. (98.9%) [MN181168] | Potato root | ||||||||||||||||
| OTU_5 | Gammaproteobacteria | Pseudomonas fluorescens (99%) [CP054128] | Soil | ||||||||||||||||
| OTU_7 | Alphaproteobacteria | Uncultured bacterium (99.6%) [JN038230] | Soil | ||||||||||||||||
| OTU_9 | Gammaproteobacteria | Marinobacter algicola (98.6%) [MK493604] | Algal culture | ||||||||||||||||
| OTU_10 | Bacteroidetes | Hymenobacter sp. (98.9%) [MH549147] | Plant | ||||||||||||||||
| OTU_13 | Alphaproteobacteria | Methylobacterium sp. (99.3%) [MN989088] | Leaves | ||||||||||||||||
| OTU_15 | Alphaproteobacteria | Novosphingobium lentum (99.1%) [AB682668] | Culture strain | ||||||||||||||||
| OTU_22 | Bacteroidetes | Gracilimonas halophila (98.8%) [NR_158001] | Water | ||||||||||||||||
| OTU_28 | Alphaproteobacteria | Phenylobacterium sp. (98.4%) [MF101711] | Hot springs | ||||||||||||||||
| OTU_37 | Bacteroidetes | Gracilimonas sp. (99%) [KJ206435] | Saltern | ||||||||||||||||
| OTU_38 | Alphaproteobacteria | Uncultured bacterium (98.6%0 [EU440697] | Soil | ||||||||||||||||
| OTU_50 | Gammaproteobacteria | Pseudomonas sp. (99.1%) [CP053697] | Grassland | ||||||||||||||||
| OTU_54 | Gammaproteobacteria | Lysobacter sp. (99.3%) [KX230693] | Soil | ||||||||||||||||
| OTU_61 | Bacteroidetes | Lewinella xylanilytica (98%) | Culture strain | ||||||||||||||||
| OTU_66 | Actinobacteria | Streptomyces sp. (99.1%) [MT538264] | Phosphatic sludges | ||||||||||||||||
| OTU_68 | Gammaproteobacteria | Pseudoxanthomonas sacheonensis (99.3%) [MF101054] | Culture strain | ||||||||||||||||
| OTU_93 | Alphaproteobacteria | Methylobacterium sp. (99.3%) [MN596044] | Phyllo-sphere | ||||||||||||||||
| OTU_103 | Alphaproteobacteria | Jannaschia sp. (98.8%) [FR693293] | Bryozoa | ||||||||||||||||
| OTU_133 | Gammaproteobacteria | Gilliamella apicola (98.4%) [MH782109] | Honeybee hindgut | ||||||||||||||||
| OTU_142 | Alphaproteobacteria | Methylobacterium sp. (99.3%) [MN989083] | Leaves | ||||||||||||||||
| OTU_144 | Alphaproteobacteria | Novosphingobium fluoreni (100%) [KY047400] | Seawater | ||||||||||||||||
| OTU_149 | Firmicutes | Uncultured bacterium (99.3%) [KC110920] | Soil | ||||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Genitsaris, S.; Stefanidou, N.; Leontidou, K.; Matsi, T.; Karamanoli, K.; Mellidou, I. Bacterial Communities in the Rhizosphere and Phyllosphere of Halophytes and Drought-Tolerant Plants in Mediterranean Ecosystems. Microorganisms 2020, 8, 1708. https://doi.org/10.3390/microorganisms8111708
Genitsaris S, Stefanidou N, Leontidou K, Matsi T, Karamanoli K, Mellidou I. Bacterial Communities in the Rhizosphere and Phyllosphere of Halophytes and Drought-Tolerant Plants in Mediterranean Ecosystems. Microorganisms. 2020; 8(11):1708. https://doi.org/10.3390/microorganisms8111708
Chicago/Turabian StyleGenitsaris, Savvas, Natassa Stefanidou, Kleopatra Leontidou, Theodora Matsi, Katerina Karamanoli, and Ifigeneia Mellidou. 2020. "Bacterial Communities in the Rhizosphere and Phyllosphere of Halophytes and Drought-Tolerant Plants in Mediterranean Ecosystems" Microorganisms 8, no. 11: 1708. https://doi.org/10.3390/microorganisms8111708